23份烟草品种遗传关系的SSR分析 
2.浙江大学农业与生物技术学院, 杭州, 310029
3.福建农林大学生命科学学院, 福州, 350002
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2011 年, 第 9 卷, 第 39 篇 doi: 10.5376/mpb.cn.2011.09.0039
收稿日期: 2010年10月08日 接受日期: 2011年03月28日 发表日期: 2011年04月08日
肖炳光等, 2011, 23份烟草品种遗传关系的SSR分析, 分子植物育种 Vol.9 No.39 (doi: 10.5376/mpb.cn.2011.09.0039)
本研究利用SSR标记分析了23份烟草品种的遗传关系。以8个烟草品种为材料从1 998对SSR引物中筛选出700对有稳定多态的引物,在23个烟草品种中检测出1 600个等位基因,平均每对引物检测的等位基因数为2.29个。23个烟草品种间遗传相似系数(GS)的变化范围为0.65~0.98、平均为0.79,其中烤烟品种间遗传相似系数在0.77~0.98之间、平均为0.86。我国烤烟主栽品种遗传基础非常狭窄,应充分利用净叶黄等地方品种作为亲本来拓宽育成品种的遗传基础。由于我国烤烟品种大多以美国引进品种为直接或间接亲本选育而成,故在聚类分析时与从美国引进的烤烟品种并未聚为各自类别。津巴布韦烤烟主栽品种或后备品种与我国烤烟主栽品种之间的遗传关系相对较远,我国可适当引进种植一定面积的津巴布韦烤烟品种,以在一定程度上缓解生产上种植品种单一化的矛盾。
我国是世界上最大的烟草生产国和消费国,但在烟草育种上却面临着过度依赖主体亲本、育成品种遗传基础狭窄等问题(王元英等, 1995)。多样化的种质资源及合理的亲本选配是育种计划成功的关键(Renganayaki et al., 2001),从分子水平上研究种质资源的遗传多样性及其亲缘关系,有助于客观了解种质资源的丰富程度及现有品种的遗传背景。
近年来,国内外利用分子标记技术对烟草种质资源进行了大量研究(何川生等, 2000; 杨本超等, 2005; 肖炳光, 2006; 杨友才等, 2006; 祁建民等, 2006; 肖炳光等, 2007; 杜传印等, 2008; Ren and Timko, 2001; Arslan et al., 2006; Siva et al., 2008),使用的分子标记主要是RAPD、AFLP、ISSR等随机性标记。SSR标记是利用SSR两侧特定的DNA序列来设计引物,扩增串联重复序列,根据串联重复数的不同揭示SSR的长度多态性,具有共显性、多态性高、重复性好、操作简便等特点(Moore et al., 1991; Akkaya et al., 1992; Morgante and Olivieri, 1993),受到广泛重视。Bindler等(2007)利用美国烟草基因组计划(TGI)的序列数据成功地开发了烟草SSR标记,并绘制了第一张基于SSR标记的烟草遗传图谱;随后,Moon等(2009)利用Bindler公布的SSR标记对美国的117份烤烟及720份引进种质(TIs)进行了研究。
本研究旨在利用Bindler等(2007)发表的SSR标记和我们自己开发的SSR标记,对23份烟草品种进行遗传多样性和遗传关系分析,为充分发掘利用烟草种质资源、拓宽育成品种的遗传基础等提供依据。
1结果与分析
1.1 SSR标记分析
利用8份参试烟草品种(编号为Nt1~Nt8)对Bindler等(2007)发表的278对SSR引物和我们自己开发的1720对SSR引物进行初步筛选,共得到700对具有稳定、清晰多态性条带的SSR引物(表1),其中119对为Bindler等(2007)发表的引物(记为PT系列),581对为我们开发的引物,包括65对基于公共数据库TIGR(http://www.tigr.org/tdb/)中所有烟草表达序列(EST)所设计的EST-SSR引物(记为Tep系列)和416对根据TGI公布的最新烟草基因组序列开发的Genome-SSR引物(记为Tp系列)。
利用上述700对SSR引物在参试的23份烟草品种(表2)中进行扩增(引物Tep008和Tp186的扩增结果见图1),共检测到1600个等位基因,每对引物检测的等位基因数在1~10个之间,平均为2.29个。表明我们开发的烟草SSR标记是成功的,同时也说明利用烟草自身的SSR引物对其进行基因组多态性分析是完全可行的。
.png) 表1 引物筛选结果 Table 1 Results of primer screening |
 图1 引物Tep008(A)和Tp186(B)的扩增结果 Figure 1 Result of amplification using primers Tep008 (A) and Tp186 (B) |
1.2烟草品种间遗传关系分析
由SSR数据估算出23份烟草品种间的遗传相似系数(表2),大小在0.65~0.98之间,平均为0.79,其中烤烟品种间遗传相似系数在0.77~0.98之间,平均为0.86。地方烤烟品种净叶黄与其它烤烟品种之间的遗传相似系数在0.77~0.79之间,遗传关系相对较远。烤烟品种云烟85(Nt9)和云烟87(Nt10)之间以及K326(Nt8)和K326J(Nt12)之间的遗传相似系数最高、达到0.98,由于云烟85和云烟87的亲本相同,K326J为K326变异株系选而成,因此其高遗传相似性是可以理解的。晒烟或香料烟与其他类型烟草品种间的遗传相似系数相对较低,分别为0.65~0.71和0.66~0.72,其中晒烟品种TI245(Nt21)与烤烟品种云烟85(Nt9)、云烟87(Nt10)、K326(Nt8)和K326J(Nt12)间的遗传相似系数最低,为0.65。上述结果表明,23个烟草品种间的遗传相似性总体相对较高,遗传基础比较狭窄;但不同类型的烟草品种之间仍存在一定的遗传差异。
.png) 表2 23个烟草品种间的遗传相似系数 Table 2 Genetic similarity coefficients between 23 tobacco cultivars |
从烤烟主栽品种的遗传关系来看,我国烤烟主栽品种如K326、云烟85、云烟87、云烟97、红花大金元等之间的遗传相似系数在0.81~0.98之间,尤其是K326、云烟85、云烟87、云烟97等之间的遗传相似系数在0.92~0.98之间,遗传相似度极高。津巴布韦烤烟主栽品种或后备品种如KRK26、KRK22、KRK23、T29、T66等之间的遗传相似系数在0.90~0.93之间,遗传相似度也非常高。而我国烤烟主栽品种与津巴布韦烤烟主栽品种或后备品种之间的遗传相似系数在0.80~0.89之间,遗传关系相对较远。
1.3聚类分析
为直观了解23个烟草品种间的遗传关系,利用700对SSR标记产生的遗传相似系数矩阵按UPGMA方法构建了聚类图(图2)。由图2可看出,在遗传相似系数GS=0.720处(L1),可将供试材料划分为四类:同属晒烟的Florida301(Nt1)和TI245(Nt21)分别单独聚为Ⅰ类和Ⅳ类;属于香料烟的Turkey Basma(Nt4)和Samsun(Nt5)聚为Ⅲ类;白肋烟Burley21(Nt2)和TN86(Nt3)及所有烤烟均聚为Ⅱ类。在遗传相似系数GS=0.875处(L2),可将Ⅱ类再分为6个亚类:Ⅱ1由白肋烟Burley21(Nt2)和TN86(Nt3)组成;Ⅱ2包括Hicks Broad Leafs(Nt6)、K326(Nt8)、K326J(Nt12)、MsK326J(Nt13)、云烟85(Nt9)、云烟87(Nt10)、云烟97(Nt11)和Coker371-Gold(Nt20)8份烤烟品种,均为从美国引进品种或具有美引品种亲缘的品种;Ⅱ3包括MsRW(Nt14)、KRK26(Nt15)、KRK22(Nt16)、KRK23(Nt17)、T29(Nt18)和T66(Nt19)6份烤烟品种,是从津巴布韦引进的品种或亲本材料;长脖黄(Nt23)、红花大金元(Nt7)和净叶黄(Nt22)则分别被单独聚为Ⅱ4,Ⅱ5和Ⅱ6亚类,其中净叶黄(Nt22)是从地方品种长脖黄(Nt23)中选育成的抗赤星病优良品种,红花大金元(Nt7)则是从美国引进的大金元品种变异株中选出的优良品种。
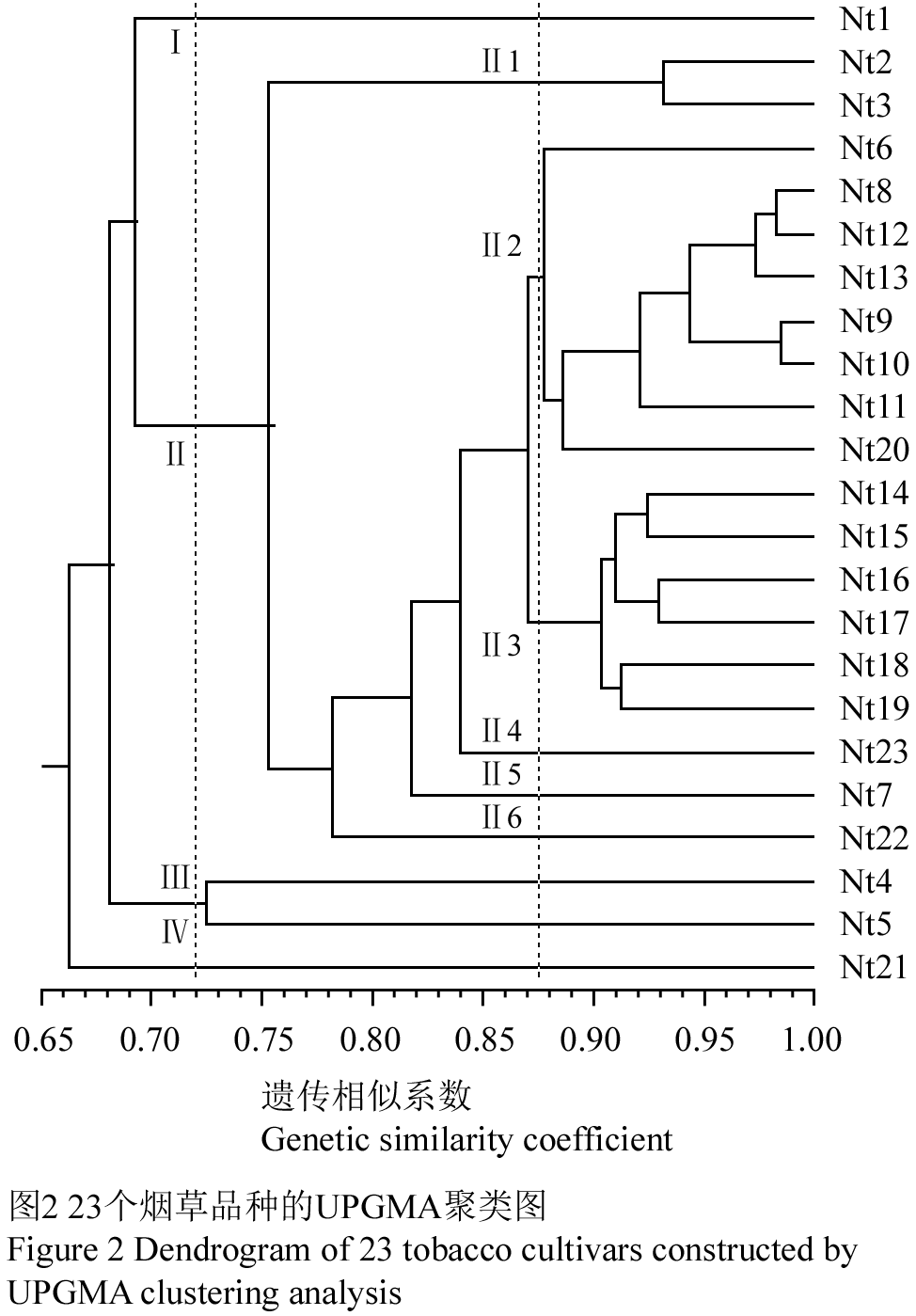 图2 23个烟草品种的UPGMA聚类图 Figure 2 Dendrogram of 23 tobacco cultivars constructed by UPGMA clustering analysis |
2讨论
2.1烟草SSR标记的遗传多态性
本研究首先利用8份烟草品种对1998对SSR引物进行筛选,获得700对多态性引物,多态性SSR引物的比例达35.04%。这一比例与Bindler等(2007)所得出的10%以内的多态率相差甚远,这是因为:(1)我们用来检测SSR标记多态性的材料为8个;(2)用于多态性筛选的1998对SSR引物中,有278对是Bindler等(2007)已在Hicks Broad Leafs和Red Russia中证实存在多态的引物,且Hicks Broad Leafs也包含在我们用来初步筛选的8个材料中;(3)我们在利用TGI发布的烟草基因组序列进行SSR引物开发时,策略上作了较大改进。在检测遗传多态性的能力方面,700个SSR标记共检测到1600个等位基因,平均每对引物可检测2.29个等位基因。而Moon等(2009)利用70个SSR引物在117份美国烤烟材料中共检测到了1031个等位基因,平均每个标记可检测到的等位基因数是14.7个,约是我们的6.14倍,这可能是因为Moon等(2009)采用了具有高灵敏度的荧光毛细电泳检测技术,而我们则是采用分辨率较低的6%非变性聚丙烯酰胺凝胶电泳检测技术。
2.2烟草品种间遗传多样性与亲缘关系
SSR分子标记聚类结果(图3)表明,烟草栽培品种间亲缘关系的远近与人为划分的烟草类型有一定的相关性。如在遗传相似系数GS=0.720处(L1),香料烟品种Turkey Basma(Nt4)和Samsun(Nt5)聚为一类,晒烟品种Florida301(Nt1)和TI245(Nt21)各自聚为一类、且与其它烟草类型分开;在遗传相似系数GS=0.875处(L1),白肋烟品种Burley21(Nt2)和TN86(Nt3)聚为同一亚类。这一结论与此前利用ISSR分析的结果比较吻合(肖炳光等, 2007)。
我国选育的烤烟品种与从美国引进的烤烟品种之间的遗传差异并未因地理来源的不同而呈明显的区别,它们聚为同一亚类、遗传关系较近,主要是由于我国烤烟品种大多以美引品种为直接或间接亲本选育而成,如云烟85(Nt9)、云烟87(Nt10)和云烟97(Nt11)的亲源中都有K326(Nt8)。Arslan等(2006)利用RAPD标记对22个来自土耳其和美国的烤烟品种的遗传多样性分析结果也表明,具有相同加工品质特性而地理来源不同的烤烟品种归为一类,与本研究的结论类似。究其原因,可能是当前的栽培品种为了迎合消费者的需求进行了严格的定向选择,而使许多非育种目标的多样化性状丢失,导致了遗传多样性降低。
我国烤烟主栽品种遗传基础极其狭窄,应充分利用净叶黄(Nt22)等地方品种作为亲本来拓宽育成品种的遗传基础。参试的津巴布韦烤烟主栽品种或后备品种间遗传相似度也非常高,其中KRK26(Nt15)的亲本之一是津巴布韦国内自主创制的种质MsRW(Nt14),推测其它几个品种的亲本之一也可能含MsRW(Nt14)或与MsRW(Nt14)亲缘较近。因我国烤烟主栽品种与津巴布韦烤烟主栽品种或后备品种聚为不同的亚类,遗传关系相对较远,从田间表现看差异也较明显,因此可适当引进种植一定面积的津巴布韦烤烟品种,以在一定程度上缓解我国烤烟生产上种植品种单一化的矛盾。
3材料与方法
3.1供试材料
参试烟草品种共23份(表3),其中烤烟17份(含津巴布韦引进品种8份)、晒烟2份、白肋烟2份和香料烟2份,均由云南省烟草农业科学研究院、中国烟草育种研究(南方)中心提供。
 表3 供试烟草品种 Table 3 Tobacco cultivars used for the experiment |
Taq Polymerase、dNTPs、100 bp DNA ladder marker 购自宝生物(大连)有限公司;丙烯酰胺和甲叉双丙烯酰胺购自USB Corporation(美国);Tis-Base、Boric Acid、EDTA-2Na和CTAB购自Amersham Biosciences(美国);其余试剂为国产。
3.2 SSR引物
所用的700对SSR引物分两部分:一部分是Bindler等(2007)发表的,共有119对;另一部分是基于TGI发布的烟草基因组序列和公共数据库TIGR(http://www.tigr.org/tdb/)提供的烟草表达序列(EST),由云南省烟草农业科学研究院[中国烟草育种研究(南方)中心]与浙江大学农业与生物技术学院联合开发的(尚未公布),共有581对。所有引物由宝生物(大连)有限公司合成。
3.3 SSR扩增
基因组DNA的提取及检测在De Riek等(2001)的方法上略作修改。
PCR扩增体系为20 μL,其中15~30 ng/μL DNA 1.5 μL、10×PCR reaction Buffer (Mg2+plus) 2 μL、25 mmol/L dNTPs 1.5 μL、10 μmol/L正反向引物各1.5 μL、rTaq 0.75 U。PCR扩增在Bio-RAD Master Cycler C1000上进行,先在94℃下预变性5min;再进行30个循环的变性(94℃, 30s)、退火(退火温度随引物的不同而改变,30s)、延伸(72℃, 30s)步骤;然后在72℃下继续延伸5~7min;最后在4℃下保存。
扩增产物加1/6体积的6×Loading Buffer,取2.5 μL利用6%的非变性聚丙烯酰胺凝胶在DYY-12型电泳仪(北京六一厂)上电泳分离,然后参照Bassam等(1991)的方法进行银染检测。
3.4数据分析
扩增产物按同一迁移位置上条带在各个材料中的有(记为1)、无(记为0)和缺失(记为2)进行统计。参试的每两份材料间的遗传相似系数(genetic similarity coefficient, GS)用Nei-Li(1979)的公式计算。利用NTSYSpc Version 2.11软件(Rohlf等, 1998)按照类平均法(UPGMA)进行聚类分析,构建聚类图。
作者贡献
肖炳光、高玉龙是本研究的实验设计和实验研究的执行人;肖炳光完成数据分析,论文初稿的写作;吴为人参与实验设计,试验结果分析,论文修改;肖炳光是项目的构思者及负责人,指导实验设计,数据分析,论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由国家烟草专卖局科技项目(110200701023)、中国烟草总公司云南省公司科技项目(05-03, 06A03, 08A05, 09YN001和2010YN02)资助。
参考文献
Akkaya M.S., Bhagwat A.A., and Cregan P.B., 1992, Length polymorphisms of simple sequence repeat DNA in soybean, Genetics, 132: 1131-1139 PMid:1459432 PMCid:1205234
Arslan B. and Okumus A., 2006, Genetic and Geographic Polymorphism of Cultivated Tobaccos (Nicotiana tabacum L.) in Turkey, Russian Journal of Genetics, 42(6): 667-671 doi:10.1134/S1022795406060123
Bassam B.J., Caetano-Anollés G., and Gresshoff P.M., 1991, Fast and sensitive silver staining of DNA in polyacrylamide gels, Analytical Biochemistry, 196: 80-83 doi:10.1016/0003-2697(91)90120-I
Bindler G., van der Hoeven R., Gunduz I., Plieske J., Ganal M., Rossi L., Gadani F., and Donini P., 2007, A microsatellite marker based linkage map of tobacco, Theor. Appl. Genet., 114: 341-349 doi:10.1007/s00122-006-0437-5 PMid:17115128
De Riek J.E., Calsyn I., Everaert E., Van Bockstaele and De Loose M., 2001, AFLP based alternatives for the assessment of distinctness, uniformity and stability of sugar beet varieties, Theor. Appl. Genet., 103: 1254-1265 doi:10.1007/s001220100710
Du C.Y., Wang Y.J., Li S.S., Li C.B., Liu H.X., and Tian J.C., 2008, AFLP Analysis of Genetic Relationship of 39 Flue-Cured Tobacco Cultivars, Zhongguo Nongye Kexue (Scientia Agricultura Sinica), 41(9): 2741-2747 (杜传印, 王玉军, 李斯深, 李常保, 刘洪祥, 田纪春, 2008, 39个烤烟种质亲缘关系的AFLP分析, 中国农业科学, 41(9): 2741-2747)
He C.S., He X.J., Li T.F., Xu M.L., and Xu J.M., 2000, Cluster analysis of flue-cured tobacco germplasm, Zhongguo Nongye Kexue (Scientia Agricultura Sinica), 33(3): 14-18(何川生, 何兴金, 李天飞, 许美玲, 许介眉, 2000, 烤烟品种资源的聚类分析, 中国农业科学, 33(3): 14-18)
Moon H.S., Nifong J.M., Nicholson J.S., Heineman A., Lion K., van der Hoeven R., Hayes A.J. and Lewis R.S., 2009, Microsatellite-based Analysis of Tobacco (Nicotiana tabacum L.) Genetic Resources, Crop Science, 49: 2149-2159 doi:10.2135/cropsci2009.01.0024
Moore S.S., Sargeant L.L., King T.J., Mattick J.S., Georges M., and Hetzel D.J.S., 1991, The conservation of dinucleotide microsatellites among mammalian genomes allows the use of heterologous PCR primer pairs in closely related species, Genomics, 10: 654–660 doi:10.1016/0888-7543(91)90448-N
Morgante M, and Olivieri A.M., 1993, PCR-amplified microsatellites as markers in plant genetics, The Plant Journal, 3(1): 175-182 doi:10.1111/j.1365-313X.1993.tb00020.x PMid:8401603 doi:10.1046/j.1365-313X.1993.t01-9-00999.x PMid:8401603
Nei M., and Li W., 1979, Mathematical model for studying genetic variation in terms of restriction endonucleases. Proceedings of the National Academy of Sciences of the United States of America, 76: 5269-5273 doi:10.1073/pnas.76.10.5269
Qi J.M., Wang T., Chen S.H., Zhou D.X., Fang P.P., Tao A.F., Liang J.X., and Wu W.R, 2006, Genetic diversity and genetic relatives analysis of tobacco germplasm based on ISSR, Zuowu Xuebao (Acta Agronomica Sinica), 32:373-378 (祁建民, 王 涛, 陈顺辉, 周东新, 方平平, 陶爱芬, 梁景霞, 吴为人, 2006, 部分烟草种质遗传多样性与亲缘关系的ISSR标记分析, 作物学报, 32: 373-378)
Ren N., and Timko M.P., 2001, AFLP analysis of genetic polymorphism and evolutionary relationships among cultivated and wild Nicotiana species. Genome, 44: 559-571 doi:10.1139/gen-44-4-559 PMid:11550889
Renganayaki K., Read J.C., and Fritz A.K., 2001, Genetic diversity among Texas bluegrass genotypes (Poa arachnifera Torr.) revealed by AFLP and RAPD markers, Theor. Appl. Genet., 102: 1037-1045 doi:10.1007/s001220000521
Rohlf F.J., 1998, NTSYSpc numerical taxonomy and multivariate analysis system, Version2.0 Exeter Software, Setauket New York, USA
Siva R.K., Sheshumadhav M. and Murthy T.G.K., 2008, Molecular diversity in the genus Nicotiana as revealed by randomly amplified polymorphic DNA, Physiology and Molecular Biology of Plants, 14(4): 377-382 doi:10.1007/s12298-008-0037-8
Wang Y.Y., and Zhou J., 1995, Parentage analysis of major tobacco varieties and tobacco breeding in America and China, Zhongguo Yancao Xuebao (Acta Tabaccaria Sinica), 2(3): 11-22 (王元英, 周 健, 1995, 中美主要烟草品种亲源分析与烟草育种, 中国烟草学报, 2(3): 11-22)
Xiao B.G., 2006, Assesment of genetic relationships between the flue-cured tobacco varieties by RAPD and ISSR markers, Wuhan Zhiwuxue Yanjiu (Journal of Wuhan Botanical Research), 24: 392-396 (肖炳光, 2006, 利用RAPD和ISSR标记分析烤烟品种间遗传关系, 武汉植物学研究, 24: 392-396)
Xiao B.G., and Yang B.C, 2007, Assessment of Genetic Diversity among Tobacco Germplasms by ISSR Markers, Zhongguo Nongye Kexue (Sientia Agricultura Sinica), 40(10): 2153-2161 (肖炳光, 杨本超, 利用ISSR标记分析烟草种质的遗传多样性, 中国农业科学, 40(10): 2153-2161)
Yang B.C., Xiao B.G., Chen X.J., and Shi C.H., 2005, Genetic diversitiy of flue-cured tobacco varieties based on ISSR marker, Yichuan (Hereditas(Beijing)), 27:753-758 (杨本超, 肖炳光, 陈学军, 石春海, 2005, 基于ISSR标记的烤烟种质遗传多样性研究, 遗传, 27: 753-758)
Yang Y.C., Zhou Q.M., and Yin H.Q., 2006, Analysis of genetic diversity in tobacco germplasm by RAPDs and ALFPs, Nongye Shengwu Jishu Xuebao (Journal of Agricultural Biotechnology), 14: 585-593 (杨友才, 周清明, 尹晗琪, 2006, 利用RAPD和AFLP标记分析烟草种质资源的遗传多样性, 农业生物技术学报, 14: 585-593)